

DISCLAIMER: The blog below is generally created from the research listed in the “Referenced Material” section. This is not my research. All credit is given to these studies and researchers who are helping beekeepers better understand how we can improve our management practices.
Introduction
Honey bee physiology follows an annual cycle, with winter bees living ten times longer than summer bees but annually we continue to see losses particularly at the end of the overwintering period. Several factors arising during the year can contribute to this. For example, climate warming or cooling impacts the lifespan in flies, fish, bees, and rodents. Other factors related to local climate changes include the quality of forage, beekeeper management practices, and pesticide exposure from land use. Although we can try to avoid some of these factors, no effective methods to mitigate the impacts of local climate changes and prevent bee winter mortality have been reported. To advance in this direction, it is necessary to clarify the general processes occurring in late summer and fall that are involved in the transition from short-lived to long-lived workers, as these processes may be disrupted by local changes. Some key triggers influencing the transition of insects from summer to winter physiology include photoperiod, ambient temperature, brood microclimate, availability of pollen, and some yet-to-be-identified factors, which obviously have a particular influence on overwintering physiological state, and also linked to aging and subsequent winter loss in bees.
Long-lived (diutinus) Phenotype
Honey bee colonies adapted to temperate climates undergo four general stages during their yearly life cycle. In the spring, worker numbers expand exponentially as colonies forage and consume abundant resources. Colonies reaching critical mass enter a colony-level reproductive phase wherein new colonies establish through swarming. Colonies then hoard resources necessary for winter survival, including extensive honey stores to fuel colony thermoregulation. Depletion of pollen stores essential for brood rearing initiates a physiological transition of young workers into a distinct long-lived (diutinus) phenotype. The diutinus phenotype acts as a storage vessel for fats and proteins destined to nourish the first spring cohort. This colony-level overwintering strategy depends largely on the production and conservation of the yolk precursor vitellogenin (Vg), a glycol-lipoprotein highly expressed in adult diutinus workers. Diutinus bees accumulate Vg, which extends their lifespan and improves their tolerance to starvation, disease, and oxidative stress. The evolution of this life history strategy has allowed honey bee colonies to endure seasonal changes in forage availability and inclement weather by storing simple sugars in the hive as an external energy source and fats and proteins, in the form of vitellogenin (Vg), within the bodies of diutinus workers. While workers during the growth phase live an average of 21 days, diutinus workers can live 150+ days in cold climates.
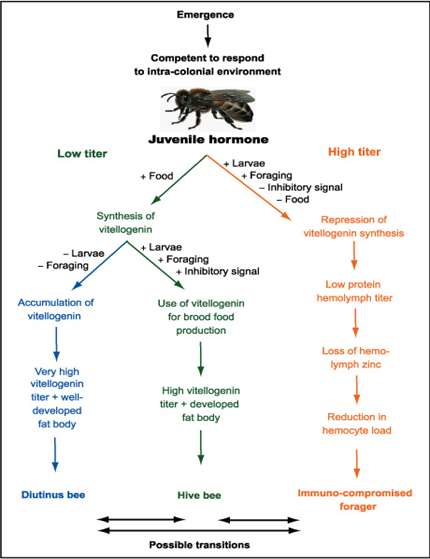
Temporal Caste and Vitellogenin
Aging and longevity are complex life history traits that are influenced by both genes and environment and exhibit significant phenotypic plasticity (change in physiology) in honey bees where different castes can have very different life spans. For example, workers are conditionally sterile and display very different aging patterns depending on which temporal caste they belong to (hive bee, forager, or a long-lived caste capable of surviving for several months on honey alone). It’s been determined that the yolk protein vitellogenin appears to play a key role in the regulatory circuitry that controls this variation.
Down-regulation of vitellogenin in foragers
Pollen is a scarce commodity, and letting foragers contain as little as possible of the major storage protein when they perish in the field may contribute positively to the reproductive output of the colony by lowering the demand for pollen foraging. It’s believed that this is the major reason for the capacity of juvenile hormone to repress vitellogenin synthesis in honeybee workers.
This explanation is in accordance with the claim that age-based division of labor, with the performance of risky tasks delayed until late in life by workers with depleted nutrient stores, may have evolved as an energy-saving mechanism in insect colonies. However, the explanation also extends this claim by showing that pathways connected to the regulation of aging can actively control this depletion of nutrient stores. Furthermore, the concerted down-regulation of the forager immune system fits very well into this explanatory picture, as it probably contributes to further nutrient deprivation as well as to a lowered nutrient demand because of reduced maintenance costs. Therefore, it seems fair to conclude that aging in honeybees is not merely a collection of nonadaptive deleterious events that happen in the shadow of natural selection, but that natural selection has shaped the aging pattern to come under strict regulatory control that ensures improved allocation of resources at the colony level.
Creation of the diutinus worker caste
A non-conception use of vitellogenin for brood production, and the down- instead of up-regulation by juvenile hormone, can be interpreted as the signature of group-level adaptation. This adaptation has probably been driven by habitat-independent, colony-level selection pressures for optimization of division of labor regimens as well as nectar and pollen collection and use. Therefore, one would expect that honeybees in both temperate and subtropical climates would share these traits. Available data on temporal vitellogenin concentrations in European and some African subspecies do indeed suggest that this is the case.
In contrast, East African lowland workers, which inhabit tropical and subtropical climates, become only slightly more long-lived, if at all, during periods of unfavorable ambient conditions and reduced brood rearing. This indicates that no strong selection pressure exists for developing a long-lived worker bee caste in major parts of Africa. On the basis of the hypothesis that vitellogenin functions as a life span-promoting storage protein in the honeybee, we would thus expect that African bees should have a substantially lower hemolymph vitellogenin titer than do European bees during periods when no broods are being produced. This appears to be the case. Taken together, these data strongly suggest that the emergence of a diutinus worker caste is an adaptive response to a more seasonal and homogeneous environment. It is difficult to envisage a simpler proximate solution than an enhanced capacity for synthesis and storage of vitellogenin.
Hypertrophied (overly developed) Hypopharyngeal Glands
The known physiological markers of winter bees’ physiology include hypertrophied hypopharyngeal glands (HG) compared to newly emerged and forager summer bees. This is important because the enlarged HGs of overwintering honey bees secrete more royal jelly to adequately feed the first larvae after diapause (superorganism hibernation). Thus, the nutrition of winter bees includes both the bee’s own nutritional needs and the nutritional needs of larvae cared for by nurse bees, which seem to stimulate each other. Another marker is an enlarged fat body compared to summer bees, resulting from the accumulation of nutrients, particularly lipids. These lipids support the production of hormones and molecular markers, as well as the synthesis of vitellogenin (vg) protein, which is later transported to the HG as a source of amino acids after diapause ends. Based on this, larger acini in the HG and higher lipid content in the fat body of winter bees may indicate a comparatively better physiological state.
Gut Microbiome and Southern Climates
Mounting evidence indicates that the honey bee hindgut microbiome has a major effect on central host metabolism, increasing health and longevity. Indeed, honey bees that lack a hindgut microbiota and consume pollen have significantly reduced Vg expression, suggesting that the microbiome is critical to central host metabolism.
Honey bee overwintering health is essential to meet the demands of spring pollination. Managed honey bee colonies are overwintered in a variety of climates and increasing rates of winter colony loss have prompted investigations into overwintering management, including indoor climate-controlled overwintering. Central to colony health, the worker hindgut gut microbiota has been largely ignored in this context but some research has found that worker longevity over winter in southern climates was much lower than that recorded for northern climates. Compared to cold indoor wintering, collective microbiota changes in the southern outdoor climate suggest compromised host physiology. Fungal abundance increased by two orders of magnitude in southern climate hindguts and was positively correlated with non-core, likely opportunistic bacteria.
Brood Temperatures
Studies have reported that short-lived summer bees tend to live longer after developing at a lower temperature (32°C/89°F). It is hypothesized that declining brood temperature from a shrinking colony size in the fall may be another trigger that affects physiological changes associated with extended longevity (long-live) in winter bees compared to summer bees. Alternatively, results have also demonstrated that constant high daytime (diurnal) temperatures negatively impact colony growth in summer and fall such as reduced brood rearing, smaller worker populations, and increased brood temperature regulation demands that might be associated with late dry summers and low foraging dearth periods.
Notably during winter temperatures conditions, bees raised at higher temperatures exhibited distinct physiological states in bees at the end of diapause (colony hibernation) in February compared to those raised at lower temperatures also suggesting that brood raised at lower temperatures in the early spring have more physiological changes associated with winter bees. This could likely be a colony evolutionary survival behavior to assure the colony has enough long-lived nurse bees later in the spring when temperatures and foraging become more appropriate for colony increase.
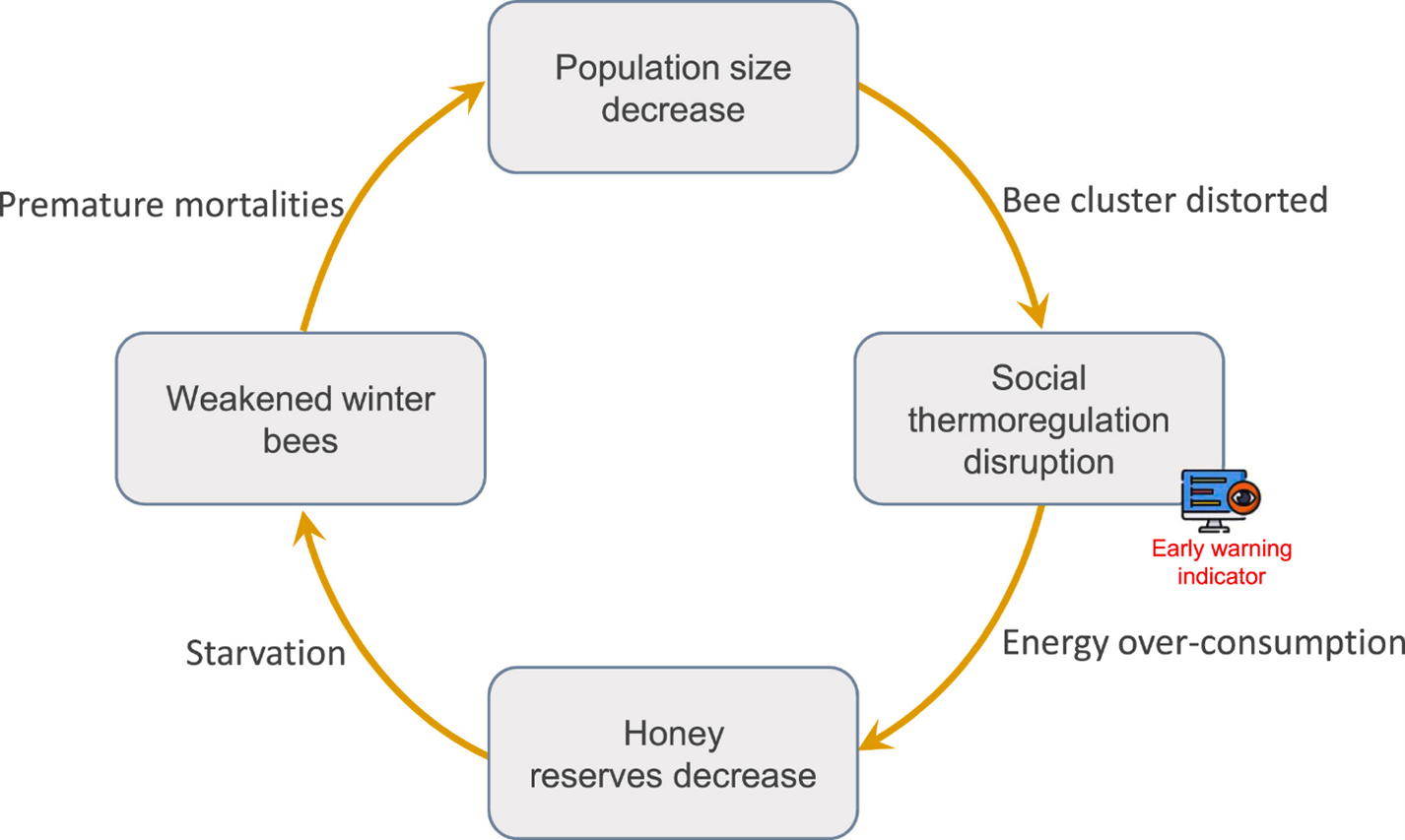
Stressors and Mortality
Identifying causal links between traits provides clues to the mechanisms of mortality. It’s suggested that population size, social thermoregulation and honey reserves are integrative traits that accumulate disturbances from individual traits and are thus better predictors of mortality while overwintering. Among these predictive traits, social thermoregulation was identified as a good candidate as an early warning indicator. Moreover, it’s suggested that the concept of a feedback loop mechanism of colony mortality for which the overwintering mechanism is centered on is the colony’s ability to optimize its social thermoregulation to limit energy loss and maintain its population of winter bees until spring. Directly or indirectly, stress factors can disrupt this mechanism, driving the colony into a feedback loop of deterioration until a critical threshold is reached, triggering first individual and ultimately colony mortality.
However, the chain of causal links is not restricted to individual traits and can spill over onto colony traits. This is the case for disturbance of bee performance, impacting both population size and social thermoregulation; and bee lifespan impacting population size. At the colony level, disturbance of population size whether through loss or increased brooding can affect social thermoregulation and honey reserves. Energy loss due to the increased effort of social thermoregulation affects honey reserves directly by triggering overconsumption. The effects of the colony’s traits both converge on honey reserves, which could be the most integrative colony-level trait. In these causal links, colony traits seem more integrative than individual traits, as they accumulate disturbances from each other and all the individual traits. So, it was identified that population size, social thermoregulation, and honey reserves together are good predictors of overwintering failure.
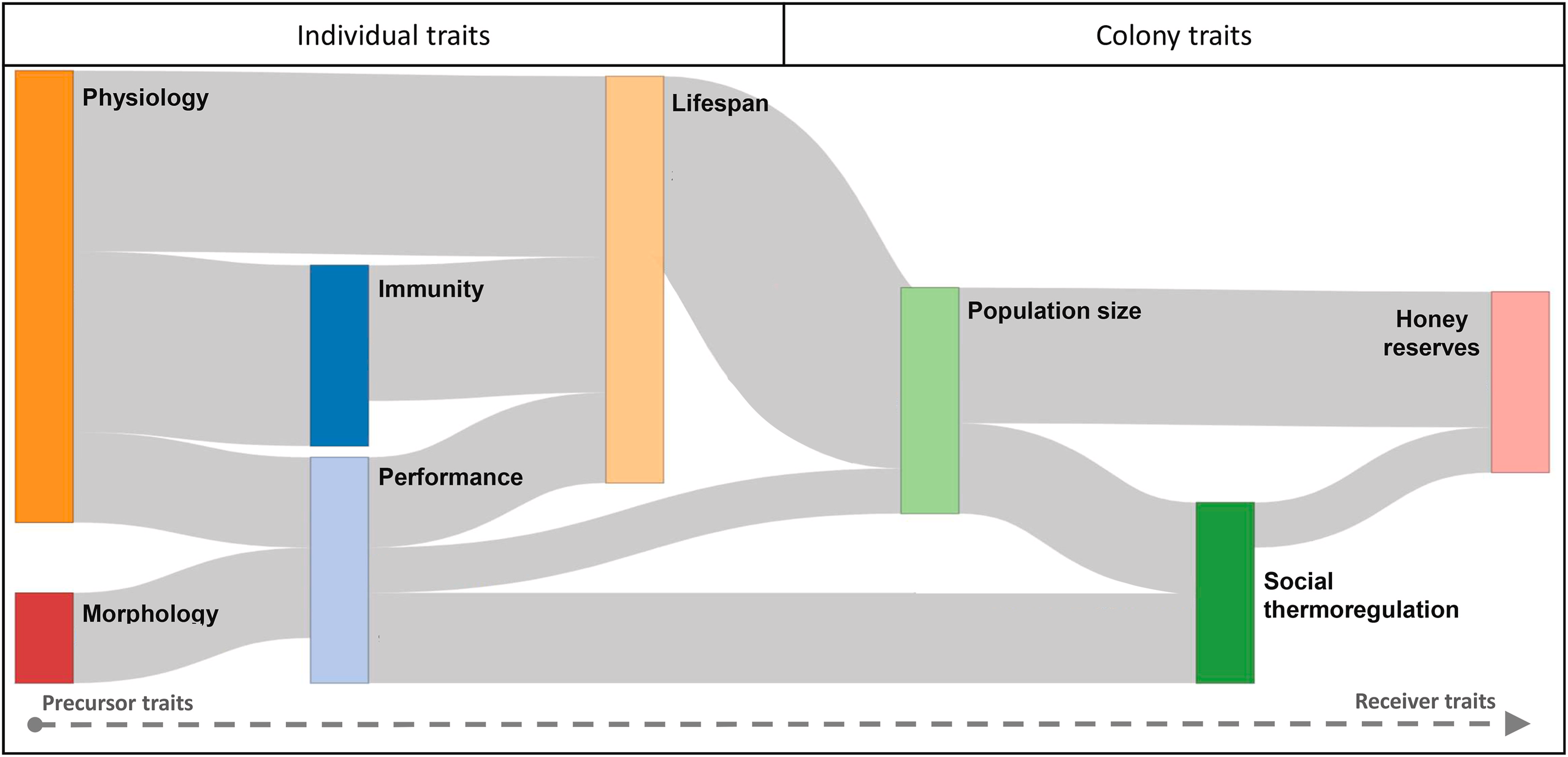
[The width of the links and the size of the biological traits are proportional to the number of studies concerned. The position of the traits is determined by calculating the number of causal effects of which it is the precursor relative to the number of causal effects it has received.]
As we see in the graph above, the causal links found between all traits for the most studied stressors (i.e. V. destructor and pesticides) give insights into potential mechanisms leading to colony mortality. For instance, adult winter bees exposed to pesticides (neonicotinoids, fungicides, or herbicides) show reduced immunity and thus increased parasite and pathogen levels such as the Varroa mite. The Varroa-infested colony begins overwintering with reduced honey reserves and population size. This population includes infested winter bees that are morphologically, physiologically, and immunologically weakened, leading to a decrease in their lifespan and an increase in their probability of dying before the end of winter. In addition, pesticides can increase the level of damaged mitochondrial DNA in the thorax of exposed honey bees, which can also represent accelerated aging and could lead to a drop in energy production and, thus, a failure of thermoregulation. Coupled with the small population, thermoregulation is disturbed, and the internal temperature is lower and/or less stable. The lower environmental temperature increases pesticide toxicity but also metabolic rate of behaviors, such as grooming. This may explain why colonies with low populations show higher consumption per individual and higher individual mortality. Over winter the colony weakens and if the population falls below a critical threshold, social thermoregulation may fail due to an insufficient number of winter bees in the colony.
Reference Materials
- The effect of seasonal temperatures on the physiology of the overwintered honey bee
- Overwintering Honey Bee Colonies: Effect of Worker Age and Climate on the Hindgut Microbiota
- How stressors disrupt honey bee biological traits and overwintering mechanisms
- Epigenetic Modifications May Regulate the Activation of the Hypopharyngeal Gland of Honeybees (Apis Mellifera) During Winter
- Effects of Wintering Environment and Parasite–Pathogen Interactions on Honey Bee Colony Loss in North Temperate Regions
- The effect of seasonal temperatures on the physiology of the overwintered honey bee
- Epigenetic Regulation of Aging in Honeybee Workers
- DNA methylation affects the lifespan of honey bee (Apis mellifera L.) workers
- Long-lived winter bees provide clues about ageing
- The rise and fall of major royal jelly proteins during a honeybee (Apis mellifera) workers’ life
- Nutritional Physiology and Ecology of Honey Bees
- More than royal food – Major royal jelly protein genes in sexuals and workers of the honeybee Apis mellifera
- Origin and function of the major royal jelly proteins of the honeybee (Apis mellifera) as members of the yellow gene family
- Colony-level pesticide exposure affects honey bee (Apis mellifera L.)royal jelly production and nutritional composition
- Oxalic Acid Treatment: Short-Term Effects on Enzyme Activities, Vitellogenin Content, and Residual Oxalic Acid Content in House Bees, Apis mellifera L.
- Queen cells acceptance rate and royal jelly production in worker honey bees of two Apis mellifera races
- Darwinian Beekeeping: An Evolutionary Approach to Apiculture