

DISCLAIMER: The blog below is generally created from the research listed in the “Referenced Material” section. This is not my research. All credit is given to these studies and researchers who are helping beekeepers better understand how we can improve our management practices.
Introduction
Below is a summary of the recent research on swarming and topics surrounding its timing and identification. Research articles are always listed at the bottom of the blog for further details. Just below are two links which contain information gleaned from the research and organized into an approximate execution timeline. Consider downloading these two resources and referencing them as you read the blog.
Remember like most things with honey bees, location is extremely important, and swarming is no different. Your area, your temperatures, your forage sources, your winter conditions all play an important part in trying to determine when swarms will happen. The downloadable timeline is an approximation for Southwest Ohio conditions. Those more north will likely be delayed by a couple weeks and those more south will be a few weeks early. Knowing what your forecasted temperatures will be and keeping an eye on your trees and early blooms will help you dial in the timeline for your area.
Fissioning
Honey bees live in colonies that reproduce by fissioning. When a colony divides itself, approximately two thirds of the workers along with the (old) mother queen, leave the hive as a swarm to found a new colony elsewhere. The rest of the workers, and a (new) daughter queen, stay behind and inherit the old nest. In cold, temperate regions, this ephemeral process of colony multiplication typically occurs only once per year and takes less than 20 minutes, making it a hard-to-study phenomenon. Resent work though has shown that signals used by honey bee colonies to initiate the departure of a swarm from its nest find that the piping signal, and the buzz-run signal, are the key signals used to initiate the swarm’s departure. The identity of the individuals that performed these signals that trigger the swarm’s exodus have also been determined and we now know that knowledgeable nest-site scouts are the producers of the signals that trigger this sudden departure. Its also interesting to know that honey bee swarms compete for, and defend, potential nest sites during their house-hunting process. While doing this they use various levels of aggression depending on the number of nest-site scouts from each swarm present at the nest site at the same time.
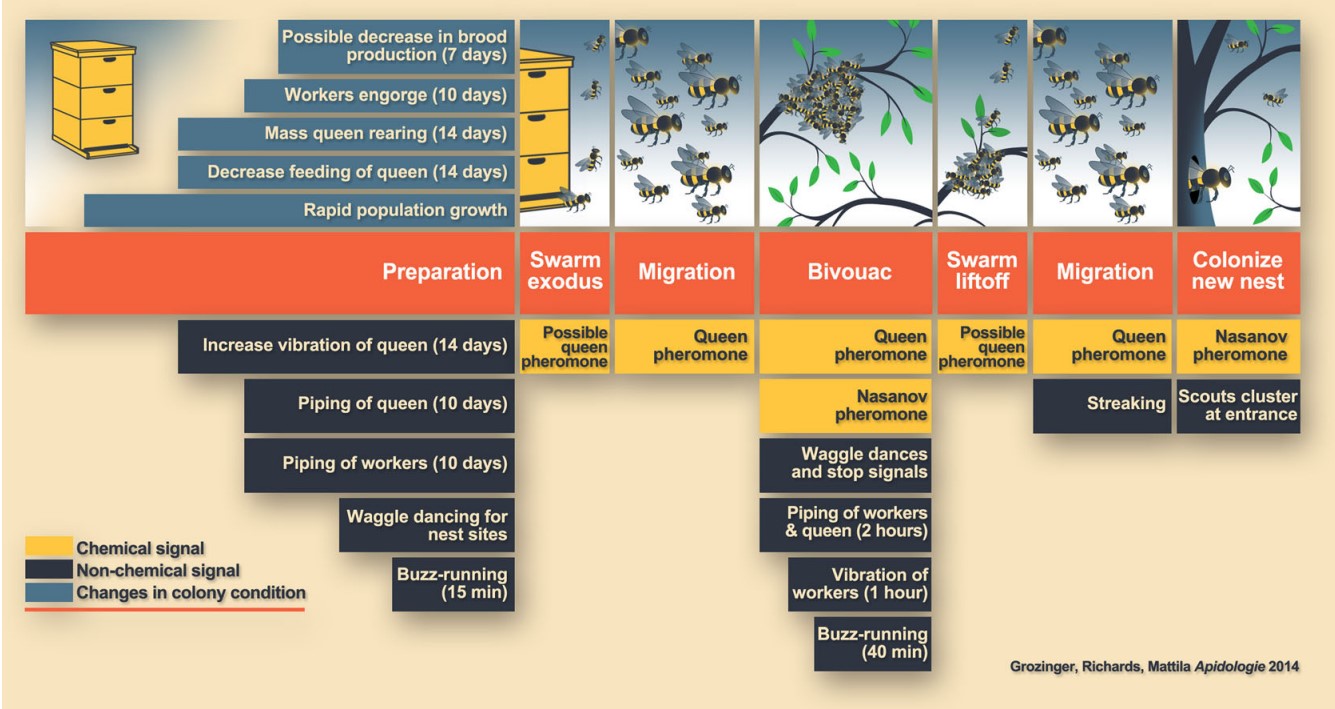
Swarming Triggers
Reproductive swarming in honey bees occurs via a series of distinct phases (shown above in orange and depicted graphically). The activities of colony members during these phases and their transitions are coordinated by nonchemical (highlighted above in black) and chemical signals (highlighted above in yellow) from queens and workers. In cases where signals coordinate swarm movement (i.e., exodus or liftoff), the periods of time prior to movement during which the signals are produced are called out. It’s hypothesized that volatile queen pheromones are involved in swarm exodus and liftoff (designated as “possible queen pheromone”). Also, “rapid population growth” prior to swarming leads to other colony conditions that may trigger swarm preparation, including increased colony size and congestion, congestion of the brood nest, and decreased concentration and/or transmission of queen pheromone. As an example, if the hive has a large population and all the frames are fully drawn, then there’s a good chance you’re about to lose bees. Other cues may include a high drone population, forager bees using the entire entrance width, queen cells, festooning bees, and large amounts of honey stored above and in the brood.
Drone Production
Honeybees are an excellent model system for examining how trade-offs shape reproductive timing in organisms with seasonal environments. Honeybee colonies reproduce two ways: producing swarms comprising a queen and thousands of workers or producing males (drones). There is an energetic trade-off between producing workers, which contribute to colony growth, and drones, which contribute only to reproduction. The timing of drone production therefore determines both the drones’ likelihood of mating and when colonies reach sufficient size to swarm. It was found that the optimal behavior for each colony is to produce all drones prior to swarming, an impossible solution on a population scale because queens and drones would never co-occur. Reproductive timing is therefore not solely determined by energetic trade-offs but by the game theoretic problem (probable decisions or strategies) of coordinating the production of reproductives among colonies.
Honey bee societies comprise three castes: workers, queens, and drones. Investment in each of these castes can conveniently represent the different methods of allocating limited resources toward different life history requirements. Workers contribute to colony growth and survival by performing all foraging, brood care, and defense. The queen’s sole job is to lay eggs; the colony rears new queens only when the colony is ready to reproduce or when the original queen requires replacement. Drones are the male reproductives of a honey bee colony. The drones consume colony resources and perform no work (some research indicates they play a key role in the thermodynamics of the nest); their traditional sole purpose is to pass on the colony’s genes by mating with queens from other colonies. Thus, workers typically eject drones from the colony in early fall once they have little chance of mating.
Following currently observed patterns for the timing of swarm and drone production, the optimal response for any individual colony would be to segregate its investment in male and female reproductives in overtime, with males being produced first and queens second. The optimal colony-level solution would not be evolutionary stable because drones and queen would never co-occur, and all colonies would have zero mating success if every colony adopted it. Selection to optimize the timing of reproduction in temperate honey bees must therefore be constrained by the need to coordinate the production of reproductives with other colonies in the population. This is accomplished by colonies maintaining drones throughout late spring and summer before and after swarming occurs.
Scouts
Recently it was discovered that a small minority of individuals in a honey bee colony—an oligarchy—decides when to trigger the departure of a swarm from its hive. The departure process begins with some bees producing the worker-piping signal (the primer for departure). Worker-piping signal, which involves a bee pressing her thorax against other bees while producing a high-frequency (200–250 Hz) vibration of her flight muscles, primes the swarm bees for flight. This is followed by these bees then producing the buzz-run signal (the releaser for departure). This minority are nest-site scouts that search for potential nest cavities prior to the departure of the swarm from its hive. Scouts, older experienced workers, also seem to have the colony job of finding new food/foraging sources.
Indications are that nest-site scouts in honeybee colonies that are preparing to swarm can start the house-hunting process before these colonies leave their hives. Also, that when predeparture nest-site scouts are interested in a potential homesite, they can return to their hive and recruit nestmates to that site by performing waggle dances. Furthermore, it was determined that the predeparture nest-site scouts are the sole producers of the worker-piping signal and that they are the first producers of the buzz-run signal. The control of the departure of a honeybee swarm from its hive shows how a small minority of well-informed individuals in a large social insect colony can make important decisions about when a colony should take action.
Departure
The observed patterns of signal production for initiating the departure of a swarm from its nest are nearly identical to those reported for priming and triggering the take-off of a swarm from its bivouac site, when it begins its flight to a new nest site. During both processes, the first signal produced by bees is the piping signal, which evidently stimulates bees to warm their flight muscles in preparation for rapid flight. It is likely that the piping signal also serves as a primer for swarm exodus, stimulating the bees to warm up in preparation for their explosive departure from the nest. The evidence that piping actually causes bees to warm up, and is not simply correlated with their warming up, comes from Seeley and Tautz (2001). When they prevented a small group of bees in a swarm from getting piped prior to swarm take-off, they found that these bees did not warm themselves and fell to the ground when the rest of the swarm bees launched into flight. The second signal that is used in both swarm departure contexts is the buzz-run. Buzz-runners begin to appear about 10 min before a swarm’s exodus from the nest, and they appear in high numbers 5 – 10 min before a swarm’s take-off from the bivouac site. Evidently, the small minority of scouts produce the buzz-run function as the “releaser” of departure in both contexts. In was reported that bees standing motionless in a small group are stimulated to activity when a buzz-running scout pushes through them.
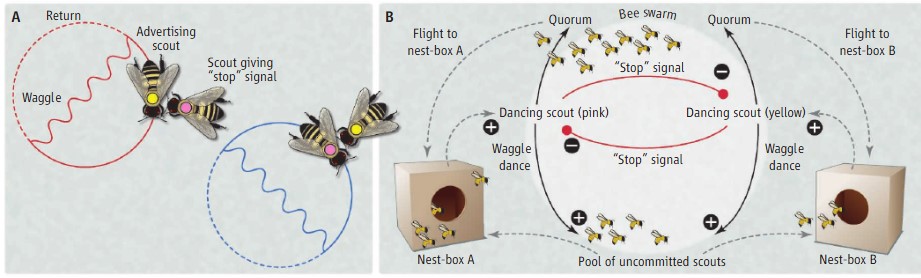
Decision Making
For a honeybee swarm of potentially thousands of individuals, choosing a home is a momentous decision. Failing to choose a single location may cause the swarm to split and the queen to be lost; choosing poorly may limit the swarm’s growth or expose it to freezing temperatures during the winter. Studies over the past 60 years have shown that honeybee swarms use quorum sensing, a form of decentralized decision-making, to choose a suitable nest site. It’s also been shown that an inhibitory signal between bees advocating different locations allows them to make a decision even when potential nest sites are equally favorable.
At the right time some of the most experienced workers leave to locate suitable nest sites. Upon their return, these scouts advertise potential locations and their qualities by performing a waggle dance. During the dance, the scout walks straight across the bees, making side-to-side waggles of her body. She then stops, turns left or right, and walks a semicircular return path to her starting point. The waggle run’s duration and orientation encode the length and the angle of the outward flight, respectively, whereas the number of dance circuits encodes the quality of the potential nest site. Waggle dances recruit additional scouts to a site until a quorum number is reached and the swarm prepares to move to its new home.
It was found that when a scout bee returns to the swarm cluster and advertises a potential nest site with a waggle dance, she tunes the strength of her dance in relation to the quality of her site: the better the site, the stronger the dance. A dancing bee tunes her dance strength by adjusting the number of waggle-runs/dance, and she adjusts the number of waggle-runs/dance by changing both the duration and the rate of her waggle-run production. Moreover, we found that a dancing bee changes the rate of her waggle-run production by changing the mean duration of the return-phase portion of her dance circuits. Differences in return-phase duration underlie the impression that dances differ in liveliness.
Bivouac
Once the bees in a swarm have left their hive and have settled into a temporary cluster hanging from a tree branch (bivouac), they show two markedly different patterns of behavior. The large majority of the swarm bees are relatively inactive and serve as food reservoirs that keep the swarm supplied with energy. A small minority (only about 5%) are active and serve as nest-site scouts that inspect potential nest cavities. The collective decision of where to move is reached through a process of competition among scouts visiting different sites, with scouts from the highest quality site producing the strongest waggle dances and thus attracting the strongest support for their site. When a quorum of nest-site scouts has built up at one of the sites, and thus a new home has been chosen, this oligarchy of nest-site scouts starts producing the worker-piping signal to prime the inactive bees in the bivouac swarm for flight. When all the swarm bees have warmed their flight muscles to a flight-ready temperature (95°F/35°C), the nest-site scouts begin producing the buzz-run signal to release the liftoff of the swarm. This whole process can happen over a day or two.
Acoustics and Vibrations
Honey bee colonies form a complex system and are known to have a large range of signals and cues for communication amongst single individuals and between single individuals and the whole hive. Whereas cues are unintentionally left by the bees, signals are an active way of information transfer. Such signals include the widely known waggle dance in which foragers communicate the direction and distance of a profitable food source, the tremble dance which aims to recruits more nectar-storing workers and many others. As colonies are self-organized in a bottom-up matter, local and global communication is crucial to ensure high efficiency of all relevant tasks. Signals can be of different nature in honeybee colonies. Olfactory signals, such as the queen pheromone which indicates queen presence to other hive members, are usually slowly released and are often present prolonged amounts of time before they are either renewed or disappear. Physical signals such as the stop signal involve bodily contact between individuals and are unsuitable for spreading wide-range messages. Vibroacoustic signals, on the other hand, are more commonly used for direct and ‘urgent’ signals about the present state of individuals or the colony and are often used in conjunction with physical contact. They can be caused by honeybees contracting their thoracic wing muscles and pressing their vibrating bodies against the wax or wooden parts of the hive or another bee. This can cause substrate-borne vibrations and result in acoustic signals. An example is the commonly used ‘whooping’ signal, previously known as ‘begging’ or ‘stop’ signal. It has a fundamental frequency of about 355 Hz, lasts for 60 ms and is transmitted on the comb as vibrations caused by the contraction of the flight muscles. Moreover, bees can also generate airborne sounds with their wings which are commonly used as signals during the waggle dance and round dance. There is no information in the literature about the resonant frequencies of the respective hive surfaces, however, bees show great sensitivity to frequencies of 280 to 350 Hz, suggesting that this is the resonant frequency of the antennal flagellum and thus a main frequency used in the conduction of the above-mentioned signals.
Tooting and Quacking
The virgin queen (not the old queen) is known for producing the substrate-borne ‘piping’ signal, which is made up of two components: ‘tooting’ and ‘quacking’. The signal is made by newly emerged queens as a sign of her presence and was thought to challenge other queens in the colony. If more than one queen is being reared, individuals that are still in capped queen cells may respond to the tooting with a quacking sound. Upon making this sound, workers can decide to protect the responding queen and feed her through a small slit in the cap or may not react to it and let the emerged queen find and kill the other queen(s). However, more recent research suggests that the piping signal serves as information to the workers about the number of capped and released queens. According to this information they can then coordinate the release of capped queens in order to decrease the rate of competition.
However, these signals do not occur later throughout the queen bee’s life (apart from in preparation for swarming) and are thus not a good indicator of queen presence for the beekeeper in the long-term. Instead, for the beekeeper the sound profile of a whole colony is a better indicator of queen status. For example, after a queen is removed a ‘warble’ signal can be detected; a sound with a frequency of 225–285 Hz and a maximum amplitude of 3 dB. It was found to develop within the first five hours after queen bee removal, before it turns into a roaring sound at 180 Hz and roughly 20 dB accompanied by vicious behaviors by female workers.
Splits, Emergency Cells, and Comb Age
If a colony is dequeened by splitting or other methods in the spring the colony will begin emergency cell creation within 24-48hrs. A study on the impact of comb age on the number of emergency queen cells and ripped queen cells was conducted comparing the morphometric and reproductive characteristics of queens reared in combs aged 1, 2, 3, and 4 years. The highest number of emergency queen cells was constructed in combs aged 1 year on the 2nd day after dequeening. No more queen cells were constructed after the 5th day of dequeening. The highest numbers of successful queen cells and emerged queens were obtained from the new combs. The amount of royal jelly (RJ)/queen cell, the weight of the newly emerged queen, and the queen cell size significantly decreased with increasing the age of the comb. Compared with the newly emerged queens from combs aged 1 year, the queens from combs aged 3 and 4 years exhibited significantly lower values for antenna length, mandibular gland area, forewing area, number of hamuli, area of the 3rd and 4th abdominal tergites, abdomen length, number of ovarioles/ovary, ovariole length and diameter, and spermathecal size. Significant positive correlations between the queen cell size and RJ yield/queen cell, queen body weight, and all queen characteristics except for the hindwing area were found. It can be concluded that the number of reared queens and their morphometrics and reproductive characteristics were significantly dependent on the age of the combs.
Reference Materials
- From molecules to societies: mechanisms regulating swarming behavior in honey bees (Apis spp.)
- A modeling approach to swarming in honey bees (Apis mellifera)
- Colony Fissioning In Honey Bees: How Is Swarm Departure Triggered And What Determines Who Leaves?
- Coordination Between the Sexes Constrains the Optimization of Reproductive Timing in Honey Bee Colonies
- An oligarchy of nest-site scouts triggers a honeybee swarm’s departure from the hive
- How Honeybees Break a Decision-Making Deadlock
- Nest-site selection in honey bees: How well do swarms implement the “best-of-N” decision rule?
- Acoustic and vibration monitoring of honeybee colonies for beekeeping-relevant aspects of presence of queen bee and swarming
- The prediction of swarming in honeybee colonies using vibrational spectra
- Identification of the honey bee swarming process by analysing the time course of hive vibrations
- The temperature increase at one position in the colony can predict honey bee swarming (Apis cerana)
- Emergency queen cell production in the honey bee colony
- Worker regulation of emergency queen rearing in honey bee colonies and the resultant variation in queen quality
- Comb age significantly influences the emergency queen rearing, morphometric and reproductive characteristics of the queens
- Honey bees consider larval nutritional status rather than genetic relatedness when selecting larvae for emergency queen rearing
- Honey bee pheromones: common scents